Выделение различных фаз адаптации организма спортсмена к высокой физиче-ской нагрузке
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Введение |
3 |
|
Материал исследования и предварительная обработка
данных |
5 |
|
Индивидуальный норматив |
8 |
|
«Зонная» модель |
13 |
|
Идентификация зон |
15 |
|
Интерпретация модели |
18 |
|
Заключение |
19 |
|
Литература |
19 |
Введение
Адаптационные свойства
организма человека определяют возможности организма и допустимую
продолжительность работы в предельных режимах. Данная работа является
продолжением исследования, посвященного моделированию процессов адаптации организма футболиста высокого класса к
высоким и предельным физическим нагрузкам [1]. Для спортсменов высокого уровня
по сравнению с физически малоактивными людьми характерны отличия по многим
физиологическим показателям (частота сердечных сокращений, артериальное
давление, объем циркулирующей крови, температура тела при выполнении тяжелой
работы, pH крови во время предельных нагрузок и др.). Такова
специфика спортивной медицины, требующая особых подходов и методов исследования.
Генерация силы и мощности во
время мышечного сокращения осуществляется за счет энергии, выделяющейся при
распаде АТФ. Для нормального функционирования мышцы концентрация АТФ в ней
должна находиться в пределах от 2 до 5 мМ [2, 3]. Ограниченные запасы АТФ в
мышце (до 5 мМ) позволяют выполнить от 4 до 6 повторений максимальных усилий
подряд или поддержать максимальную мощность упражнения в течение 1-2 с. Поэтому
для обеспечения непрерывной сократительной деятельности мышцы АТФ в ней должна
восстанавливаться с такой же скоростью, с какой она расщепляется в процессе
сокращения.
Ресинтез АТФ при мышечной деятельности
возможен как в ходе реакций, идущих без потребления кислорода (анаэробный
механизм), так и за счет окислительных превращений в клетках (аэробный
механизм). Обычно ресинтез АТФ происходит в основном путем аэробных
превращений. При напряженной мышечной
деятельности доставка кислорода к мышцам затруднена и в тканях усиливаются анаэробные процессы ресинтеза
АТФ. В скелетных мышцах человека возможны три вида анаэробных процессов ресинтеза
АТФ: креатинфосфокиназная реакция, миокиназная реакция; гликолиз (называемый
также лактатным анаэробным процессом, он сопровождается образованием молочной кислоты,
лактата, с которым сопряжено развитие утомления в работающих мышцах). Образования
молочной кислоты в первых двух процессах не происходит, их называют алактатными
анаэробными механизмами. Анаэробные процессы имеют преимущество при выполнении
кратковременных упражнений высокой интенсивности, а аэробные – при длительной
работе умеренной интенсивности.
Для определения состояния здоровья спортсмена
и уровня его работоспособности применяются процедуры нагрузочного тестирования.
Материалом данной работы послужили результаты нагрузочных тестов 133
футболистов Российской премьер-лиги. При игре в футбол задействованы все
основные способы энергообеспечения – аэробный, анаэробный алактатный и
анаэробный лактатный. В силу большой длительности футбольного матча, основным
является аэробный механизм [4].
Интегральным показателем аэробного метаболизма является потребление кислорода в
единицу времени, т.е. динамика потребления кислорода.
Цель данного исследования - создание моделей
процессов адаптации организма спортсмена к физической нагрузке для решения
основных задач спортивной физиологии:
1) Определение максимальных
адаптационных возможностей организма;
2) Выявления факторов,
лимитирующих адаптационные возможности организма;
3) Определение соотношения
аэробного и анаэробного механизмов
энергообеспечения организма. Признаки
изменения этого соотношения (перехода). Возможности прогнозирования переходов,
их физиологические предвестники.
Решение этих задач должно помочь при поиске
перспективных спортсменов и выборе
адекватной спортивной специализации для них, определении функциональной
готовности спортсмена к соревнованиям, планировании его тренировок, а также при
выработке стратегии и тактики во время соревнования.
В данном исследовании использовались идеи
динамики открытых и сложных систем [5], методы аппроксимации и
непараметрической статистики [6, 7].
Традиционно для динамики потребления
кислорода в начале выполнения упражнения с постоянной мощностью используют
полиэкспоненциальную модель, представляющую интенсивность потребления кислорода
суммой экспонент [8]. Это есть следствие моделирования динамики потребления
кислорода системами линейных дифференциальных уравнений с постоянными коэффициентами.
Определение коэффициентов этих моделей
представляет собой трудную техническую задачу. Прямое их измерение часто
практически невозможно, а оценка по
регистрируемым траекториям показателей требует достаточно хорошего начального
приближения, подбор которого при большом количестве параметров модели является
тяжелой задачей.
Кроме того, уравнения с постоянными
коэффициентами в ряде случаев дают излишне грубую модель. Например, часто
считают постоянной активность ферментов, влияющую на скорость метаболических
реакций. Однако активность многих ферментов резко снижается при снижении
внутриклеточного pH, например, при активной работе анаэробных лактатных
механизмов [2, 3] или при изменении температуры [2]. Заметим, что в работающих
мышцах отмечается повышение температуры вследствие тепловыделения в результате
метаболических реакций.
Таким образом, популярные ныне имитационные
модели адаптации, основанные на системах линейных дифференциальных уравнений с
постоянными коэффициентами, имеют достаточно узкую область применимости. Это
приводит к трудностям распространения полиэкспоненциальной модели, отмеченным в
[8], на область высоких нагрузок, наиболее полно отражающих адаптационные
возможности организма и представляющих наибольший интерес для данного
исследования.
Материал исследования и предварительная
обработка данных
Стендовому тестированию были
подвергнуты футболисты-мужчины Российской премьер-лиги (133 человека). Во время
тестирования испытуемые бегут в «гору» по дорожке, установленной под небольшим
углом к горизонту. Угол наклона дорожки можно менять, не прерывая тестирование.
Скорость бега принудительная, задается врачом. Продолжительность испытания не
превышала 20 минут. В процессе тестирования
регистрировали следующие показатели: легочную вентиляцию, частоту дыхания,
потребление кислорода, выделение углекислого газа и частоту сердечных сокращений.
Испытание проводили с
использованием эргоспирометрической установки Oxycon Alpha
(Рис. 1) фирмы Jaeger (Германия).
Рис. 1. Эргоспирометрическая установка Oxycon Alpha
Антропометрические и
возрастные данные протестированных футболистов представлены в Табл. 1 (min
– минимум, q1 – нижний квартиль, med – медиана, q2 – верхний квартиль, max
– максимум, av – среднее, stdev – стандартное отклонение).
Табл. 1.
Антропометрические и возрастные данные протестированных футболистов
|
|
min |
q1 |
med |
q2 |
max |
av |
stdev |
|
рост [см] |
168 |
178 |
181 |
185 |
190 |
180.8 |
5.7 |
|
масса [кг] |
61 |
72 |
76 |
80 |
91 |
75.7 |
6.4 |
|
возраст [лет] |
16 |
19 |
22 |
26 |
33 |
22.6 |
4.4 |
Нагрузка спортсмена в этом
упражнении определяется двумя параметрами: скоростью бега и уровнем наклона
дорожки (G[%]), который связан с углом наклона дорожки
α соотношением: α = arctan(G/100).
Нами использовался следующий протокол проведения испытаний (Рис. 2):
1)
Разминка: бег со скоростью 5 км/час при уровне подъема дорожки 0.5 в
течение 1.5 минут;
2)
Основная нагрузка. Бег со ступенчатым увеличением нагрузки и постоянной
длительностью ступеней. Начальная скорость бега – 7 км/час, уровень подъема
дорожки 0.5; возрастание скорости бега при переходе к следующей ступени на 1.5
км/час, увеличение уровня наклона дорожки на 0.2, длительность каждой ступени 2
мин.
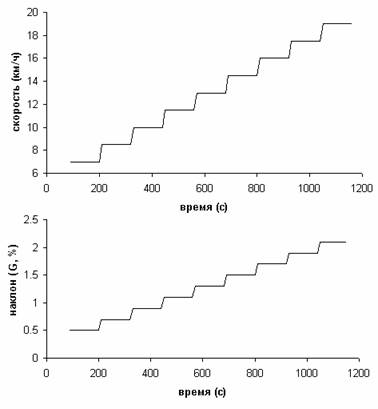
Рис. 2.
Протокол нагрузки
Тестирование
завершали при выполнении хотя бы одного из критериев [1,
9] достижения
максимального потребления кислорода – одной из основных характеристик
аэробных возможностей организма [8, 9]. Кроме того, тестирование может быть прекращено по
требованию спортсмена.
Известно, что спортсмены в
игровых видах спорта не любят монотонных физических нагрузок и часто прекращают
выполнение физической нагрузки, не достигнув своих предельных показателей. В
этом случае максимальное потребление кислорода также не достигается. Однако
некоторые параметры физического состояния спортсмена удается оценить даже в
таких ситуациях.
Все показатели
регистрируются на каждый вдох/выдох. На Рис. 3 представлена динамика
потребления кислорода в расчете на килограмм массы тела (называемого также
относительным потреблением кислорода (ОПК), [мл/(мин·кг)]) у спортсмена с
номером НОМЕР=70. По оси абсцисс отложено время [с], по оси ординат – ОПК.
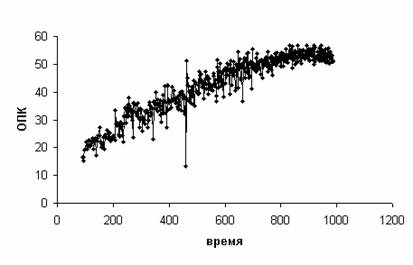
Рис. 3.
Пример динамики потребления кислорода в расчете на килограмм массы тела
Запись потребления кислорода
сопровождается помехами, включая заведомые артефакты (например, очень низкое
значение в середине записи, 459 с). Для их устранения нами использовалось
медианное сглаживание [10]. При этом для каждого сегмента значений аргумента в
качестве значения функции в центральной его точке используется медиана
распределения значений ординаты на нем. Медиана малочувствительна к изолированным
выбросам. Число точек сегмента выбирали нечетным (h точек в каждую сторону,
всего 2h+1). Для вычисления значений у концов кривых никаких специальных
приемов не применялось, поэтому h первых и h
последних измерений пропадают. При низких значениях h наблюдается остаточный шум,
а при высоких теряется много измерений в начале и конце теста. На Рис. 4
приведены результаты медианного сглаживания с различными значениями h.
Вся дальнейшая обработка проводилась при h=5.
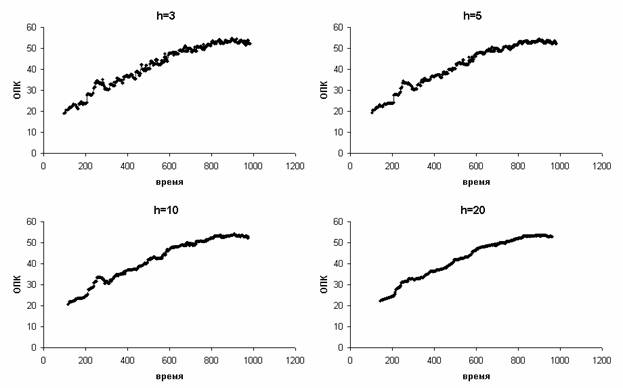
Рис. 4.
Результаты скользящего медианного сглаживания для разных значений h
Индивидуальный норматив
На Рис. 5 представлена
динамика ОПК для двух футболистов (с номерами НОМЕР=13 и НОМЕР=70). Динамика
ОПК у обоих футболистов качественно похожа, но имеет различные диапазоны
изменения по ординате.
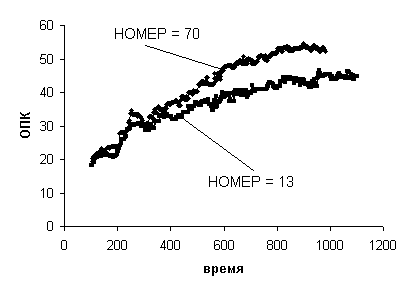
Рис. 5.
Динамика ОПК для двух футболистов
Оценим их сходство точнее.
Отложим по абсциссе значения ОПК13, а по ординате значения ОПК70
для тех же моментов времени (Рис. 6).
Точки легли вдоль прямой, соответствующей
линейной регрессии (R- коэффициент корреляции):
ОПК13 = 0.695 · ОПК70 +
6.54 (R2=0.997).
Таким образом, мы видим, что
кривые одинаковы с точностью до линейного преобразования, поэтому можно
привести ОПК70 к масштабу ОПК13. Соответствующую величину назовем
приведенным ОПК (ОПКП) и будем вычислять по формуле:
ОПКП70 = 0.695 · ОПК70
+6.54
На Рис. 7 приведена динамика
ОПКП для рассматриваемых спортсменов: здесь ОПКП13 совпадает с ОПК13.
Видно, что динамика ОПКП у обоих футболистов действительно очень похожа.
Аналогично мы можем привести
динамику ОПК всех спортсменов к масштабу ОПК спортсмена с НОМЕР=13, выбор
которого был обусловлен максимальной продолжительностью тестирования. Параметры
распределения R2 линейных регрессий, приводящих все кривые к единому
масштабу, представлены в Табл. 2.
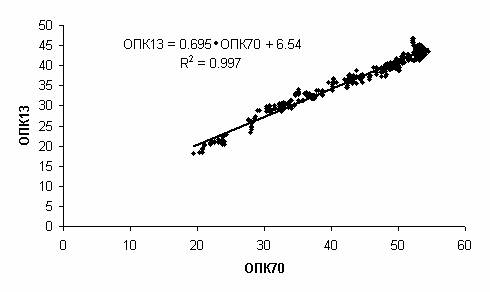
Рис. 6.
Связь ОПК двух футболистов
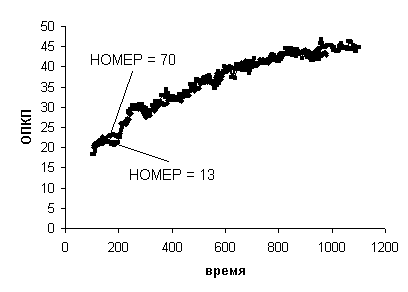
Рис. 7.
Динамика приведенного ОПК (ОПКП) двух футболистов
Табл. 2.
Распределение R2 линейных
регрессий
|
|
min |
q1 |
med |
q2 |
max |
|
R2 |
0.9806 |
0.9961 |
0.9971 |
0.9978 |
0.9988 |
Применим технологию
скользящих нормативов [6]. Скользящим нормативом случайной величины x(t) в
диапазоне t1<t<t2 будем называть распределение
F(x,t) , явно зависящее от параметра t , такое, что P(x(t)<a)=F(a,t),
t1<t<t2. Эмпирические скользящие нормативы
строят обычно на основе серии гистограмм случайной наблюдаемой величины на
последовательности смежных интервалов параметра t. Для характеристики
изучаемого процесса удобно использовать квантильные линии y(t),
проведенные через заданные квантильные точки интервальных распределений. Чаще
всего используют линии медианы и двух квартилей – верхнего и нижнего. Полосу
между заданными квантильными линиями
иногда называют нормативным коридором.
Для ОПК и ОПКП нами были построены скользящие
нормативы (с разбиением оси времени на интервалы по 10 с). На Рис. 8
представлено сравнение ширины (определяемой как разница между максимумом и
минимумом или как межквартильный интервал) нормативного коридора для ОПК и
ОПКП. Нормативный коридор для ОПКП оказался значительно уже нормативного
коридора для ОПК, что говорит о том, что по динамике ОПКП тестируемые
спортсмены больше похожи друг на друга.
Преобразовывая скользящие нормативы, построенные для
ОПКП, преобразованием обратным тому, которым производилось приведение к одному масштабу, для
каждого из тестируемых спортсменов получаем свой индивидуальный скользящий
норматив ОПК.
Поведение в индивидуальном
нормативном коридоре в каждый момент времени характеризуется квантилем qu,
которому соответствует измеренное значение ОПК. Переход к этой переменной
позволяет сравнить динамику процессов у различных спортсменов в едином
укрупненном масштабе и в дальнейшем ввести типизацию самих спортсменов по
характеру процессов, сопровождающих нагрузочное тестирование.
Поскольку значения qu в нормативных коридорах для ОПК и ОПКП совпадают,
для удобства мы определяли qu в координатах ОПКП. Для
вычисления квантиля qu мы аппроксимировали
интервальные распределения F(x(t),t)
распределением Вейбулла:
![]() , l > 0, a > 0,
, l > 0, a > 0,
параметры которого оценивались с помощью медианы med, нижнего q1 и верхнего q2 квартилей интервальных распределений [6]:
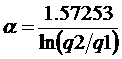 ,
,
![]() .
.
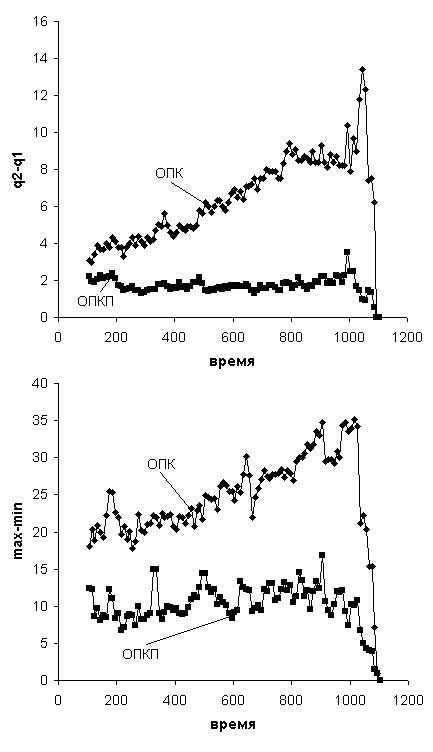
Рис. 8.
Сравнение ширины нормативного коридора для ОПК и ОПКП
На Рис. 9 представлены
эмпирическое распределение (F) значений ОПКП для всех 133
спортсменов во временном интервале 700-710с (924 точки) и аппроксимирующее его
распределение Вейбулла (F_weib).
На Рис. 10 приведена связь F и F_weib,
прямая линия соответствует линейной регрессии. Как видим, распределение
Вейбулла достаточно хорошо аппроксимирует эмпирическое распределение.
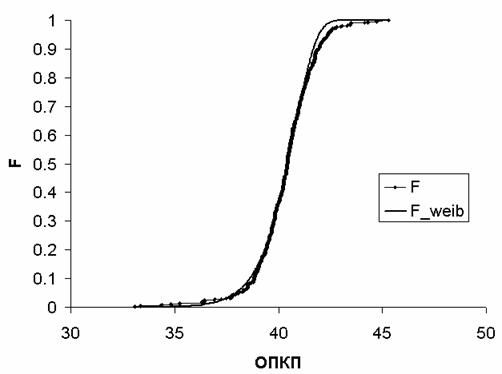
Рис. 9.
Аппроксимация эмпирического распределения распределением Вейбулла
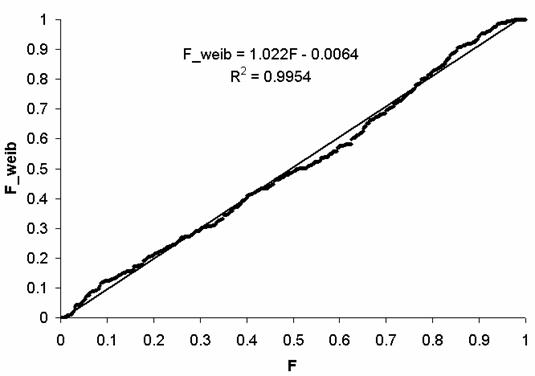
Рис. 10.
Связь эмпирического распределения и аппроксимирующего его распределения Вейбулла
На Рис. 11 приведена динамика qu для
спортсмена с НОМЕР=70. На
этом графике наблюдаются тенденции изменения qu: снижение через некоторое
время после начала теста, затем заметный рост и снова снижение в конце. Значительно
лучше эти медленные тенденции видны на Рис. 12 после медианного сглаживания по
41 точке (по 20 крайних точек пропадают).
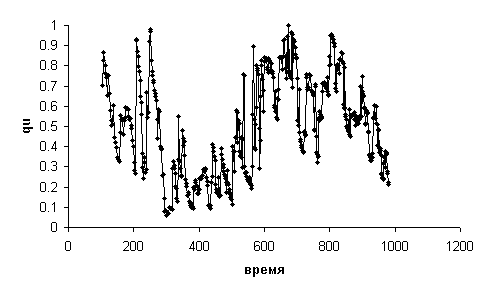
Рис. 11.
Пример динамики qu (без сглаживания)
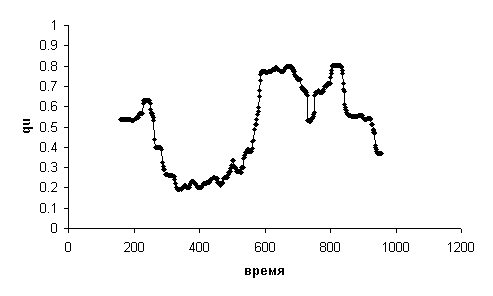
Рис. 12.
Пример динамики qu (медианное сглаживание по
41 точке)
Видно, что за время тестирования значения qu
не остаются постоянными, а достаточно сильно меняются. Характер этих изменений
(т.е. динамика qu) может быть связан с индивидуальными особенностями
организма спортсмена. Таким образом, оказывается возможным построение простого
и адекватного языка для описания динамики qu, что может помочь выделить
те факторы, которые влияют на адаптацию спортсмена к высокой физической нагрузке
в ходе тестирования.
«Зонная» модель
Для дальнейшего анализа динамику qu
удобно представить в виде монотонно неубывающей функции. Для преобразования к
такой функции можно использовать, например, интеграл с переменным верхним
пределом, который обозначим I(qu):
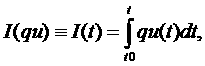
где t – время, а t0 – момент времени, которому
соответствует первое значение qu. Функция I(qu) является монотонно
неубывающей, поскольку значения qu неотрицательны.
На Рис. 13 приведен результат применения
такого преобразования функции qu, представленной на Рис. 12.
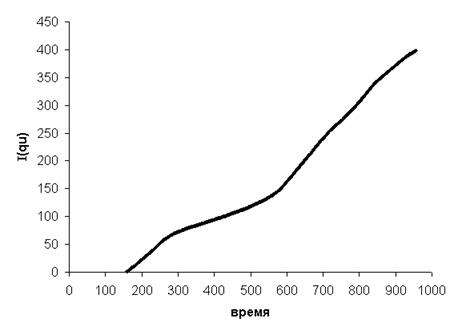
Рис. 13. Пример функции I(qu)
Эта функция неплохо приближается ломаной.
Можно выделить три зоны, в каждой из которых функция I(qu) аппроксимируется прямой
(Табл. 3, Рис. 14). В таблице каждая из линейных зон охарактеризована временем
начала и конца ([с]), и параметрами линейной аппроксимации I(qu) = a·время
+ b. Чем выше r2 для зоны, тем точнее она
соответствует линейной модели.
Каждому линейному участку функции I(qu) можно сопоставить постоянное
значение qu (Рис. 15). Таким образом, получаем «зонную»
модель: можно выделить несколько
зон, в каждой из которых динамика потребления кислорода описывается постоянным
значением qu. Такая модель не противоречит физиологии процесса адаптации
к высоким нагрузкам в виде разноуровневого многозвенного
процесса, где на разных стадиях определяющую роль могут играть разные звенья.
Далее остаются две важные задачи: идентификация зон и физиологическая
интерпретация.
Табл. 3.
Аппроксимация функции I(qu) кусочно-линейной функцией
|
Зона |
Начало, с |
Конец, с |
a, 1/с |
b |
r2 |
|
1 |
160 |
290 |
0.55 |
-87.97 |
0.998 |
|
2 |
290 |
560 |
0.24 |
-0.25 |
0.995 |
|
3 |
560 |
956 |
0.68 |
-241.94 |
0.997 |
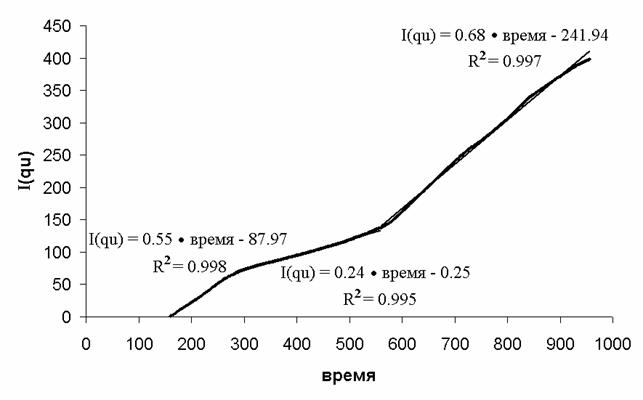
Рис. 14. Аппроксимация функции I(qu) линейными зонами
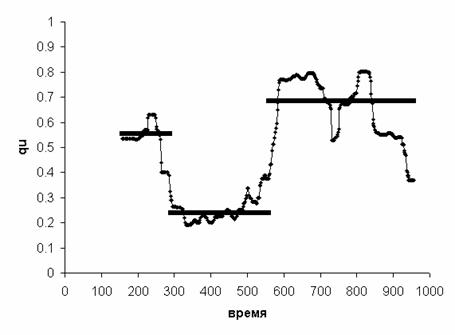
Рис. 15.
«Зонная» модель
Идентификация зон
Задача идентификации зон
сводится к аппроксимации монотонно неубывающей функции I(qu) кусочно-линейной функцией.
Для определенности будем рассматривать пример (Рис. 13), используя удовлетворительное
приближение (Табл. 3).
Сначала рассмотрим задачу нахождения границ
между зонами при известном количестве зон и их примерном расположении.
Допустим, мы хотим аппроксимировать
рассматриваемую функцию тремя линейными зонами. При этом обладаем априорной
информацией о том, что момент времени 200 с лежит в 1-ой зоне, момент времени
400 с – во второй зоне, а 800 с – в третьей. Тогда для нахождения границ между
зонами можно использовать следующий алгоритм (А1):
Шаг 1. Инициализация зон. Выбираем две точки,
ближайшие к 200 с. Относим их к 1-ой зоне. Поскольку через две точки всегда
можно провести прямую, характеризуем 1-ую зону соответствующей прямой и
значением r2=1. Аналогично инициализируем остальные зоны.
Шаг 2. Поиск вариантов
расширения зон. Каждый вариант включает в себя расширяемую зону и точку, которая может
быть включена в эту зону. Эта точка, во-первых, должна быть свободной, то есть
еще не включенной ни в одну зону, а во-вторых, соседней, то есть между этой
точкой и точками зоны не лежит никаких других точек. Таким образом, вариантов
не больше удвоенного числа зон.
Шаг 3. Выбор и реализация
варианта расширения. Расширение состоит во включении точки в зону и линейной аппроксимации
расширенной зоны методом наименьших квадратов, значением r2 которой характеризуется
реализуемый вариант. Среди всех выбирается вариант с наибольшим значением r2.
Шаг 4. Проверка на наличие
свободных точек. Если свободных точек не осталось, то выполнение алгоритма завершается.
Если еще остались, то происходит возвращение к Шагу 2. Поскольку за каждую
итерацию цикла количество свободных точек уменьшается на одну, а их количество
ограничено, то завершение работы алгоритма происходит за конечное число шагов.
В Табл. 4 приведены характеристики зон,
полученных при применении данного алгоритма.
Допустим, что в рассматриваемом нами примере мы знаем число зон – три, но
ошиблись в оценке их месторасположении и инициализировали их парами точек
вблизи моментов времени 200 с, 700 с и 800 с
(то есть по-другому инициировали 2-ую зону). Тогда получим гораздо худшие
результаты (см. Табл. 5). Таким образом, важно правильно оценить расположение
зон. Очевидно, знание количества зон необходимо для инициализации.
Описанный алгоритм естественным образом
обобщается на любое число зон, лишь бы была априорная информация, необходимая
для их инициализации. Первая и последняя точки в любом случае попадают в
крайние зоны, поэтому первую зону удобно инициализировать двумя первыми
точками, а последнюю – двумя последними точками.
Табл. 4.
Результат применения алгоритма А1 (характеристики зон)
|
Зона |
Начало, с |
Конец, с |
a, 1/с |
b |
r2 |
|
1 |
160 |
279 |
0.56 |
-89.61 |
0.999 |
|
2 |
282 |
524 |
0.23 |
3.49 |
0.998 |
|
3 |
524 |
956 |
0.68 |
-239.34 |
0.997 |
Табл. 5.
Результат применения алгоритма А1 (характеристики зон, другая инициализация)
|
Зона |
Начало, с |
Конец, с |
a, 1/с |
b |
r2 |
|
1 |
160 |
409 |
0.38 |
-48.98 |
0.953 |
|
2 |
411 |
744 |
0.55 |
-150.94 |
0.953 |
|
3 |
746 |
956 |
0.63 |
-196.93 |
0.993 |
В общем случае нет априорной информации о
числе зон и их расположении. Тогда можно аппроксимировать заданную функцию
большим числом линейных зон, чтобы получить достаточную точность приближения.
Это возможно, поскольку ломаная, соединяющая отрезками соседние точки, заведомо
дает приближение с r2=1. В отсутствие априорной
информации о положении зон естественно при инициализации зон требовать, чтобы
расстояние между любыми соседними инициализирующими парами точек было меньше
предполагаемого минимального размера линейной зоны (критерий плотности
покрытия). Итак, при достаточно большом числе зон мы можем получить высокую
точность приближения исходной функции. С другой стороны, линейных зон не должно
быть очень много, иначе теряется смысл такой аппроксимации. Более того, при
большом числе зон часто соседние зоны лежат практически на одной прямой. Тогда
их разумно объединить. Например, если r2 для объединенной зоны не
меньше минимума r2 объединяемых зон. В этом случае объединение аппроксимируется
не хуже, чем одна из зон до объединения (критерий 1). Если по этому
критерию зоны нельзя объединить, но r2 для каждой из них выше
требуемой точности, то можно предложить еще один критерий объединения зон: две
соседние зоны могут быть объединены, если r2 для объединенной зоны не
меньше заданного уровня r2 (критерий 2).
Можно предложить такой алгоритм
кусочно-линейной аппроксимации функции:
Шаг 1. Выполнение алгоритма А1, на
вход которого подается достаточно большое число зон, удовлетворяющих критерию
плотности покрытия.
Шаг 2. Объединение зон по критерию
1. Сначала объединяются зоны с максимальным результирующим r2. Шаг повторяется до тех
пор, пока есть кандидаты на объединение.
Шаг 3. Объединение зон по критерию
2. Шаг осуществляется в случае надобности при задании требуемого уровня
точности. Сначала объединяются зоны с максимальным результирующим r2. Шаг повторяется до тех
пор, пока есть кандидаты на объединение.
В Табл. 6 представлена информация о
количестве зон и их r2 (минимум, медиана и
максимум) после каждого шага алгоритма А2 при инициализации 21 зоной (разбиение
времени тестирования на 20 равных интервалов) и заданном уровне r2=0.99. Интересно, что в
результате мы получили те же 3 зоны, что и в приведенном в начале раздела удовлетворительном
приближении (Табл. 2). Характеристики полученных зон приведены в Табл. 7.
Табл. 6. Количество зон и их r2 при применении алгоритма А2
(пример)
|
Шаг |
Число зон |
r2 |
||
|
min |
med |
max |
||
|
1 |
21 |
0.99843 |
0.99976 |
1 |
|
2 |
15 |
0.99878 |
0.99976 |
0.99996 |
|
3 |
3 |
0.99608 |
0.99670 |
0.99889 |
Табл. 7.
Результат применения алгоритма А2 (характеристики зон)
|
Зона |
Начало, с |
Конец, с |
a, 1/с |
b |
r2 |
|
1 |
160 |
261 |
0.57 |
-90.45 |
0.999 |
|
2 |
264 |
494 |
0.23 |
2.88 |
0.996 |
|
3 |
495 |
956 |
0.67 |
-231.23 |
0.997 |
Результат оказался немного
хуже, чем при применении алгоритма А1 для трех зон с правильной оценкой их
положения (Табл. 4), значение r2 для 2-ой зоны оказалось
ниже. Но получен этот результат был без априорной информации о зонах, в чем и
состоит ценность предложенного нами алгоритма А2.
Интерпретация модели
В соответствии с
рассматриваемой зонной моделью динамика qu представляет собой
ступенчатую функцию (то есть в каждой из зон динамика потребления кислорода
описывается постоянным квантилем в индивидуальном нормативном коридоре). Наша
гипотеза состоит в том, что внутри разных зон разные звенья процесса адаптации
к высоким физическим нагрузкам играют определяющую роль. Особый интерес
представляют переходы от одной зоны к другой, когда в организме спортсмена
происходят изменения, «переводящие» его на другой уровень qu.
Понимание этих изменений должно позволить выяснить особенности зон. Таким
образом, задача интерпретации переходов между зонами приобретает ключевое
значение.
Переход на более высокий
уровень qu может быть связан с дополнительным ростом
потребления кислорода, что может быть обусловлено снижением эффективности использования
потребленного кислорода или дополнительными энерготратами организма (например,
при нарушении координации движений или при увеличении работы дыхательных мышц
при учащении дыхания).
Переход на более низкий
уровень qu может быть связан со снижением темпов роста
потребления кислорода вследствие исчерпания аэробных ресурсов организма или
увеличения эффективности использования потребляемого кислорода.
Более детальная
физиологическая интерпретация предложенной модели станет темой дальнейшей
работы. Для решения задачи интерпретации предполагается использовать методы
непараметрической статистики и язык логических симптомов [6].
Заключение
Разработана процедура построения
индивидуального нормативного коридора потребления кислорода в расчете на
килограмм массы тела (называемого также относительным потреблением кислорода,
ОПК). Для описания динамики показателей ОПК
нами предложена «зонная» модель, позволяющая для каждого
испытуемого выделить несколько зон, в каждой из которых потребление кислорода
находится на приблизительно постоянном уровне в индивидуальном нормативном
коридоре. Такая модель позволяет обратить внимание исследователя на видимые
стадии процесса адаптации организма к высоким физическим нагрузкам.
Кусочно-линейная
аппроксимация кривых позволяет уточнить уровни переменных и границы зон. Для
осуществления такой аппроксимации построен алгоритм, не требующий априорной
информации о зонах.
Переходы с одного уровня
показателей на другой могут быть сопряжены с изменением соотношения
аэробного и анаэробного механизмов
энергообеспечения организма. Это соотношение является важным фактором при определении
функциональной готовности спортсмена к соревнованиям, планировании его
тренировок, а также при выработке стратегии и тактики во время соревнования.
Подробная физиологическая интерпретация предложенной модели станет темой
последующих работ.
Автор выражает глубокую
благодарность Ю.Б. Котову, А.В. Гусеву, В.И. Павлову, З.Г. Орджоникидзе,
Ю.П.Попову, Г.Г. Малинецкому, В.В. Самойленко, А.В. Ермакову, Н.И. Волкову,
А.В. Подлазову за обсуждение и поддержку.
Работа частично поддержана
РФФИ. Грант 07-01-00376а.
Литература
1.
Гусев А.В., Котов Ю.Б., Орджоникидзе З.Г., Павлов В.И., Эсселевич И.А.
Исследование динамики высоких физических нагрузок с помощью компьютерного
тестирования и методов математического моделирования. – Информационные технологии и вычислительные системы
– 2007 – №1 – с.49-55
2.
Волков Н.И., Несен Э.Н., Осипенко А.А., Корсун С.Н. Биохимия мышечной деятельности. Олимпийская
литература (Киев), 2000. – 503 с.
3.
Физиология человека: Учебник для вузов физ. культуры и факультетов физ.
воспитания педагогических вузов / Под общ. ред. В.И. Тхоревского. – М.: Физкультура,
образование и наука, 2001. – 492 с.
4.
Stolen T., Chamari K.,
Castagna C., Wisloff U. Physiology of
soccer. An update. Sports Med., №35 (6) – p. 501-536.
5.
Малинецкий Г.Г. Хаос. Структуры. Вычислительный эксперимент: Введение в
нелинейную динамику. – М.: УРСС, 2000. – 256 с.
6.
Котов Ю.Б. “Новые математические подходы к задачам медицинской
диагностики” – Москва, УРСС, 2004. – 328 с.
7.
Стентон Гланц “Медико-биологическая статистика”, перевод с английского –
Москва, Практика, 1999
8.
Jones A.M. and Poole D.C.
Oxygen uptake kinetics in sport, exercise and medicine. – Routledge,
Taylor&Francis Group, London and New York, 2005.
9.
К. Wasserman, Hansen J.E., Sue D.Y. et al.
Exercise testing and interpretation. – Lippincott Williams&Wilkins, 2005.
10. Тюрин Ю.Н., Макаров А.А. Статистический анализ данных
на компьютере / Под. ред. В.Э.Фигурнова – М.: 1998. – 528 с.